Wednesday Discussion on Geometric Issues
This session was moderated by John Shareshian.
The discussion started off with some comments about Riemannian metrics
and whether the set of metrics on tree space would be useful to
study.
- Vogtmann asked if this space may be too large to study?
- Forman noted that when geometers look at the space of all Riemannian
metrics on a given space, they do it to try to find the ``best''
metric in some sense (e.g., such as one with constant curvature).
Q. Are there many useful or interesting metrics on tree space?
Q. Is the BHV metric the only one (up to scalars)
of non-positive curvature? (Bridson)
- Charney: metrics are slightly different, but they should all be
similar. If there is no reason that biologists suggest that one
is better than another, then it makes sense to choose one that has
useful properties, such as metrics of nonpositive curvature.
- Billera: for biologists, what metrics do you want to have on the
orthants?
Q. What metrics ``should'' be used on the subspace obtained by fixing
the tree type?
- St. John: uses the
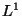 metric, though Felsenstein uses
metric, though Felsenstein uses 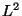 .
.
- Penny: we use because then lengths scale with time.
- Charney: since the data is not completely in tree space, maybe we
should be considering non-intrinsic metrics.
- Forman: want to choose metrics so that the statistical methods we are
using are continuous.
- Flath: perhaps some edges are more important than others?
- Penny: we are not surprised when we get edges near leaves are accurate,
but we are really interested in getting deep internal edges right.
- Penny: taxonomic studies, just care about the branch order (weight 1
on edges), but when we consider time studies, we do want the lengths.
- Forman: measuring the residuals seem to imply that embedding tree
space in some larger space and considering an extrinsic metric
would be important.
- Vogtmann noted that Felsenstein was talking about embedding
tree space in Euclidean space (using the Robinson-Foulds metric).
Q. How does the Robinson-Foulds metric compare with the BHV metric?
(Diaconis) What's the Lipshitz constant? (Bridson)
Q. Should the metric on tree space come from an intrinsic metric,
or an extrinsic metric on a larger space in which tree space is
embedded? Which is a more natural way to view tree space?
- Forman: studying the residuals seems to be very important. This
larger space is part of what you are given in the data, and should be useful.
Charney rephrased the question as:
what is the right ambient space to embed tree space? While Bridson
remarked that he viewed tree space as God-given, Forman asserted that
he views the data as God-given.
- Vert remarked that the ``average''
of trees with high likelihood may not be high
likelihood with the intrinsic metric.
Q. How do you deliver geometry to biologists? (Billera)
- Billera pointed out that interesting mathematics in biology
seems to happen by serendipity-- cases in which someone
happened to know some mathematics or knew someone who did.
- Penny: notes that there is a role for mathematics to play.
Also new majors in mathematical biology (at some schools) have arisen
that will train students to think in both disciplines.
- Diaconis notes that as a result of this conference, and ensuing
discussions, people might write an article for Science. This may bring
biologists to mathematics.
- Snel suggests advocating a notion of average,
and communicate why it is better. For instance,
articles on concatenated alignments have appeared in Science.
- Huelsenbeck: write review articles that explain things clearly.
Write for Systematics Biology--
that's where stuff on trees would appear.
- Billera asked again how one spreads mathematical knowledge in the biological
community, noting that courses offered at most institutions are
either at too low or too high a level for biologists.
Back to the
main index
for Geometric models of biological phenomena.